
A mutagénese mediada por transposões é um processo biológico que permite a inserção de genes no cromossoma do organismo hospedeiro, interrompendo ou modificando a função de um gene alvo e causando mutação. Trata-se de um processo de mutagénese mais eficiente que a mutagénese química, apresentando uma maior frequência de mutação e uma menor probabilidade de levar à morte do organismo. Outras vantagens incluem a capacidade de induzir mutações únicas, a capacidade de se incorporar marcadores de selecção na construção de uma estirpe e a capacidade de reverter os mutantes após a mutagénese. Mas também apresenta algumas desvantagens: baixa frequência de transposição e na falta de precisão dos sistemas de transposição na mutagénese.
Hitória
A mutagénese realizada por parte de transposões foi estudada pela primeira vez por Barbara McClintock em meados do século XX, durante o seu trabalho de investigação com milho, o cujo lhe garantiu um prémio Nobel. Ela trabalhou na área da Botânica durante o seu Doutoramento, tendo estudado os cromossomas do milho. Nos anos 40, McClintock encontrava-se a estudar a progenia de plantas de milho autopolinizadas e apercebeu-se que alguns cruzamentos possuíam um cromossoma 9 quebrado, faltando-lhe os telómeros. Este estudo levou à descoberta dos elementos de transposição, a partir dos quais a mutagénese assistida por transposões se desenvolveu.
Dinâmica do sistema

{kind=link}
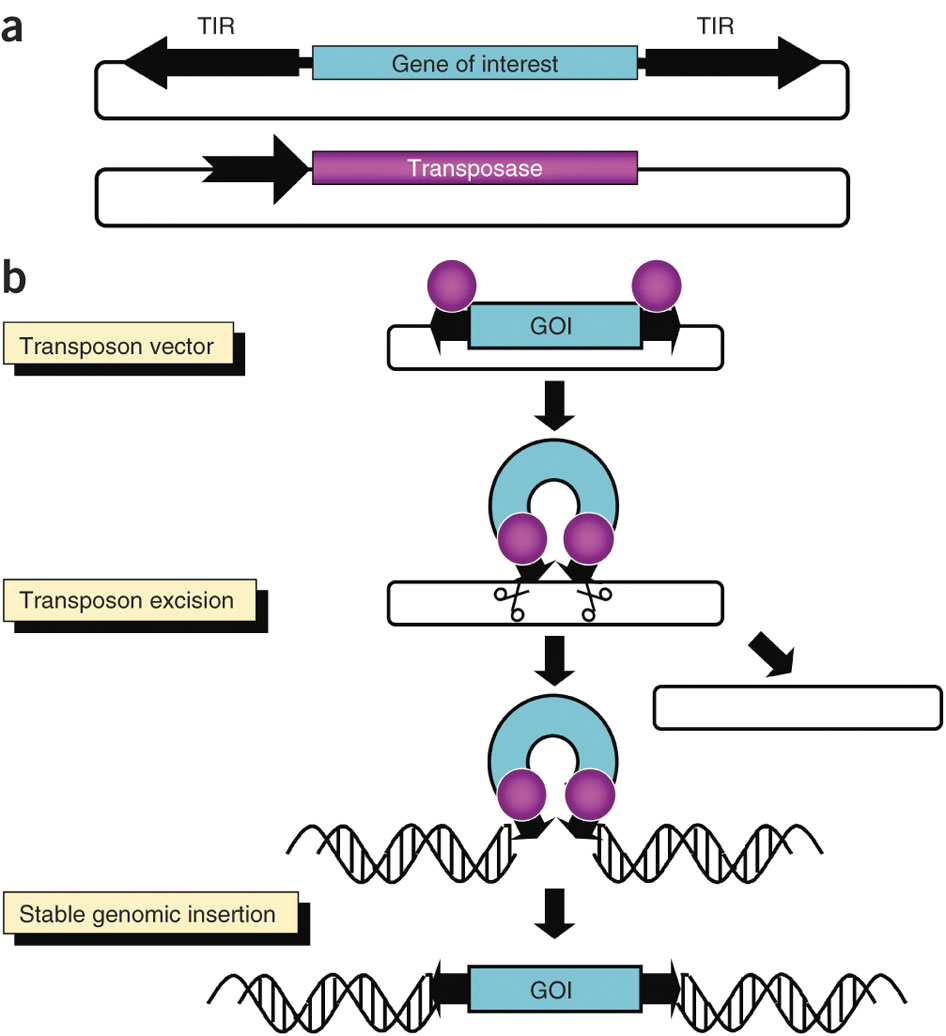
Em algumas bactérias, a mutagénese mediada por transposões é conseguida normalmente via um plasmídeo, a partir do qual o transposão é extraído e inserido no cromossoma do hospedeiro. Isto costuma requerer um conjunto de enzimas, incluindo uma transposase, a serem traduzidas. A transposase pode ser expressa tanto a partir de um outro plasmídeo, como a partir do plasmídeo que contém o transposão. Alternativamente, uma injecção de mRNA que codifica para a transposase na célula hospedeira pode induzir expressão e tradução. As primeiras versões desta técnica baseavam-se na utilização de bacteriófagos e plasmídeos conjugativos bacterianos para a inserção de sequências. Não se tratava de uma forma muito específica e era difícil a sua utilização para a incorporação de genes específicos. Uma nova versão, denominada shuttle mutagenesis usa genes específicos clonados de espécies hospedeiras para incorporar elementos génicos. Isto facilita a integração destes elementos no cromossoma e a expressão do transgene a longo prazo.
Sistema de transposição Tn5
O sistema de transposão Tn5 é um sistema modelo para o estudo de transposição e de aplicação em mutagénese. Tn5 é um transposão bacteriano em que genes (o transposão original contém genes de resistência a antibióticos) são flanqueados por duas sequências de inserção idênticas – IS50R e IS50L, correspondendo aos lados direito e esquerdo do transposão, respectivamente. O IS50R codifica para duas proteínas – Tnp e Inh. Estas duas proteínas são idênticas em sequência, excepto o facto do Inh não possuir os 55 primeiros aminoácidos da região N-terminal. A proteína Tnp corresponde a uma transposase e a Inh corresponde a um inibidor da transposase. O domínio de ligação ao DNA da Tnp reside nos primeiros 55 aminoácidos da região N-terminal, pelo que são essenciais para a função que esta desempenha. As sequências IS50R e IS50L são flanqueadas por elementos de 19 pares de bases, tanto nas extremidades internas como nas externas do transposão – IE (extremidade intrna) e OE (extremidade externa). Mutações nestas regiões resultam na impossibilidade da transposase se ligar às sequências. Esta interacção entre a transposase e estas sequências é bastante complicada, e é afectada por metilação do DNA e outros marcadores epigenéticos.
O mecanismo mais aceite para a transposição do Tn5 é o mecanismo comum para todos os sistemas de transposões. Este começa com a Tnp a ligar-se Às sequências IE e OE de cada sequência IS50. As duas extremidades ligam-se, por oligomerização do DNA e são cortadas para fora do cromossoma. Após da introdução de extremidades 5’ de 9 pares de bases nas sequências de DNA alvo, o transposão (e os genes que este contém) é inserido nessas sequências alvo, duplicando as regiões em cada extremidade do transposão. Genes de interesse podem ser introduzidos no sistema de transposão, entre as sequências IS50R e IS50L. Ao colocar o transposão sob controlo de um promotor do hospedeiro, os genes serão expressos quando introduzidos no seu cromossoma. Além dos genes de interesse, é comum introduzir-se ainda marcadores de selecção, para identificar os transformantes.
Aplicações práticas
Como resultado da capacidade da mutagénese por recurso de transposões incorporar genes em regiões alvo dos cromossomas, existem diversas aplicações associadas a este processo.
Genes de virulência de vírus e bactérias podem ser descobertos por disrupção de genes e observação do fenótipo. Isto tem grande importância para a produção de antibióticos e controlo de doenças.
Genes não essenciais podem ser identificados pela indução de mutação destes num organismo. Se após a integração homóloga do transposão num determinado gene (ou conjunto de genes) o organismo sobreviver, então o gene (ou conjunto deles) é não-essencial. Os genes mutados podem ser confirmados por PCR do DNA genómico do organismo usando primers específicos para a grelha de leitura (ORF) dos genes em questão e primers específicos para o transposão. Como os transposões podem introduzir-se em regiões não codificantes do DNA, os primers específicos para a grelha de leitura garantem avaliar se o transposão interrompeu o gene.
Genes relacionados com a oncogénese também podem ser identificados através da mutagénese e avaliação das características fenotípicas dos tumores. Dependendo do mecanismo de mutagénese e nos resultados, os genes podem ser classificados como oncogenes ou como genes supressores tumorais.