
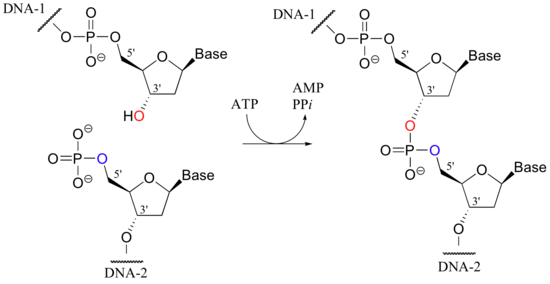
Em Biologia Molecular, ligação é a junção de dois fragmentos de ácidos nucleicos por acção de uma determinada enzima (uma ligase). Trata-se de um método de laboratório essencial em manipulação de DNA e em clonagem molecular, por fim a obter-se uma molécula de DNA recombinante (e.g. inserção de fragmentos de DNA em plasmídeos). As extremidades dos fragmentos são juntadas pela formação de ligações fosfodiéster entre o grupo hidroxilo da extremidade 3’ de um fragmento e o grupo fosfato da extremidade 5’ do outro fragmento. RNA também pode ser ligado de forma similar. Para que esta reação ocorra, é necessário adicionar-se ATP e/ou NAD+. Por norma, a reação de ligação é realizada utilizando-se a T4 DNA ligase.
Ligação de um fragmento de DNA (Insert) a um plasmídeo (Vector).
São vários os fatores que influenciam as reações de ligação, tais como a concentração da enzima e dos reagentes, a temperatura da reação e o tempo de incubação. A ligação é uma reação complicada, dado que se trata de uma reação que envolve interações inter e intra-moleculares, cujo produto corresponde a uma junção de duas moléculas de DNA diferentes. Além disso, um passo adicional de emparelhamento (annealing) é necessário para uma ligação eficiente.
A concentração do DNA pode afetar a taxa de ligação e, acima de tudo, pode determinar de a reação é intermolecular ou intramolecular. A ligação envolve a junção de extremidades de uma molécula de DNA com as de outra molécula. No entanto, cada molécula possui duas extremidades, pelo que, se estas forem compatíveis, uma molécula de DNA pode circularizar unindo ambas as suas extremidades. A elevadas concentrações de DNA, existe uma grande probabilidade de ocorrer uma ligação intermolecular, enquanto que a baixas concentrações de DNA, é mais provável ocorrer uma reação intramolecular.
A eficiência de transformação com DNA linear é muito inferior do que quando se usa DNA circular, e para que o DNA circularize, a concentração deste não deve ser demasiado elevada. Como regra geral, o DNA total não deve exceder os 10 µg / mL.
A concentração relativa dos fragmentos de DNA, bem como os seus comprimentos, são também fatores que podem influenciar se as reações intermoleculares são favorecidas, relativamente às intramoleculares.
A concentração do DNA pode ser aumentada artificialmente por adição de agentes de condensação, como cloreto de hexaaminocobalto(II) ou aminas biogénicas (e.g. espermidina), ou crowding agentes (como polietilenoglicol – PEG) que aumentam a concentração efetiva das enzimas. No entanto, é importante de se notar que aditivos, como o hexaaminocobalto(II)2+ podem levar a reações intermoleculares excessivas, resultando em concâmeros lineares em vez de DNA circular, muito mais viável para transformação com DNA plasmídico. No caso de ser necessário recorrer-se ao uso destes aditivos, é preferível o uso de PEG, dado promover tanto reações intermoleculares como reações intramoleculares.
Quanto maior a concentração de ligase, mais rápida é a taxa de ligação. A ligação de extremidades abruptas é muito menos eficiente do que a ligação de extremidades coesivas, por isso uma maior concentração de ligase é recomendada para as ligações de extremidades abruptas. Também se pode PEG em conjugação com uma elevada concentração de DNA ligase para uma ligação mais rápida (estes componentes já costumam estar presentes nos kits de ligação rápida).
Relativamente à temperatura, há duas questões que devem ser tidas em consideração nas reações de ligação. Em primeiro lugar, a temperatura óptima para a atividade da DNA ligase, que é 37ºC, e em segundo lugar, a temperatura de melting (Tm) das extremidades das moléculas de DNA a serem ligadas. A temperatura de melting é dependente do comprimento e do tipo de bases azotadas que compõe as extremidades a ligar – uma maior percentagem de guanina (G) e citosina (C) corresponde a uma maior Tm, dado estabelecerem um maior número de pontes de hidrogénio e, por isso, requererem mais energia para quebrar as ligações – com alguma contribuição por parte do grau de empacotamento das bases entre os fragmentos. Para que a ligação se proceda de forma eficiente, as extremidades devem emparelhar-se de forma estável e, na prática, a Tm das extremidades de DNA é geralmente inferior a 37ºC. A temperatura óptima para a ligação de extremidades coesivas é portanto um compromisso entre a temperatura mais adequada para a atividade enzimática e a temperatura de melting do local de ligação. No entanto, diferentes enzimas de restrição originam diferentes extremidades e a composição dessas extremidades também varia, pelo que a temperatura recomendada depende fortemente das enzimas de restrição usadas. As ligações também podem ser realizadas com recurso a extremidades compatíveis diferentes de enzimas de restrição diferentes na mesma mistura reacional, pelo que não se torna prático ajustar a temperatura para uma ligação particular. A maioria dos protocolos recomenda uma temperatura de 12-16ºC, temperatura ambiente ou então 4ºC (dependendo do protocolo).
A força iónica da solução tampão pode influenciar a ligação. O tipo de catiões presentes também pode afetar a reação. Por exemplo, um excesso de Na+ pode fazer com que o DNA fique mais rígido e promover reações intermoleculares. A elevadas concentrações de catiões monovalentes (>200 mM), a ligação pode ser quase completamente inibida. As soluções tampão padrão estão desenhadas para minimizar os efeitos iónicos.

Ligação de extremidades coesivas:
As enzimas de restrição podem gerar uma vasta gama de extremidades de DNA, mas em técnicas de clonagem, utiliza-se maioritariamente as enzimas que originam extremidades em cadeia simples com 4 bases azotadas, chamadas extremidades coesivas (com exceção da NdeI, que gera uma extremidade de 2 bases, e as que originam extremidades abruptas). Estas extremidades coesivas podem emparelhar-se com outras extremidades compatíveis e unirem-se numa ligação coesiva. Para a maioria das enzimas de restrição, a Tm das extremidades geradas ronda os 15ºC (excepto a EcoRI, que gera uma extremidade com uma Tm de 6ºC). Por questões práticas, ligações de extremidades coesivas são realizadas a 12-16ºC ou à temperatura ambiente, ou então a 4ºC por períodos mais prolongados.
Para a inserção de um fragmento de DNA num plasmídeo vetor, é preferível usar-se duas enzimas de restrição diferentes para digerir o DNA, para gerar um fragmento com extremidades incompatíveis entre si, de forma a evitar religação do vetor sem o fragmento a inserir e evitar a ligação bidirecional do fragmento. Se não for possível usar dois locais de corte diferentes, então o vetor pode ser desfosforilado para evitar um excesso de vetor religado “vazio”. Isto é feito, normalmente, recorrendo-se à fosfatase alcalina intestinal de vaca (CIAP), que remove o grupo fosfato da extremidade 5’ do DNA digerido. Mas é importante referiri que a CIAP é difícil de inativar e esta pode interferir com a reação de ligação. A CIAP não deve ser usada em excesso e só deve ser usada quando necessária.
Ligação de extremidades abruptas:
Ligações de extremidades não-coesivas, abruptas ou cegas, não envolvem emparelhamento de bases de extremidades protuberantes, pelo que não há extremidades abruptas incompatíveis. A maior vantagem de se usar enzimas de restrição que originam extremidades abruptas é que o fragmento a inserir no vetor não requere nenhum local de restrição na sua sequência, uma vez que extremidades abruptas são, por norma, originadas na amplificação por PCR. Este amplificado pode então ser diretamente ligado a um vetor digerido com uma enzima que origine uma extremidade abrupta.
A ligação de extremidades abruptas é muito menos eficiente que a de extremidades coesivas. Tipicamente, a reação é 100 vezes inferior à ligação de extremidades coesivas. Dado que não possuem extremidades protuberantes, a ligação de extremidades abruptas depende de colisões aleatórias dos fragmentos. Para compensar a falta de eficiência desta ligação, costuma usar-se uma maior concentração de DNA ligase (pelo menos 10 vezes maior do que em ligações de extremidades coesivas). A concentração de DNA também deve ser superior para aumentar o número de colisões e é recomendado prolongar-se a duração da reação.
Se ambas as extremidades do fragmento que se pretende ligar ao plasmídeo forem abruptas, então é necessário desfosforilar o vetor para minimizar religação do vetor “vazio”. è também necessário fosforilar-se o fragmento a inserir no vetor por forma a existir um grupo fosfato na extremidade 5’ que permita a ligação fosfodiéster. Normalmente recorre-se a um tratamento do fragmento com T4 polinucleótido cinase.
Ligações de extremidades não-coesivas são reversivelmente inibidas por altas concentrações de ATP.
Embora o PCR gere extremidades abruptas, é preciso ter-se em atenção que ao usar a Taq DNA Polimerase pode levar à adição de adeninas extrea na extremidade 3’ dos produtos amplificados. Estas extremidades podem ser usadas nas ligações TA, em que o vetor é tratado com terminal transferase, na presença de didesoxitimidina trifosfato (ddTTP), para gerar uma extremidade com timinas extra que irá ligar-se à extremidade com adeninas extra.
Orientações gerais para a inserção de um fragmento num plasmídeo circular:
A concentração total de DNA não deve exceder os 10 µg / mL;
O rácio molar do fragmento para o vetor deve rondar os 3:1. Um rácio muito elevado pode originar múltiplas inserções. O rácio deve ser ajustado ao comprimento do fragmento.
Problemas na ligação:
DNA danificado – sobre-exposição a radiação UV durante a preparação do DNA para ligação pode reduzir significativamente a eficiência de transformação. A adição de citidina ou guanosina ao tampão de eletroforese a uma concentração de 1 mM pode ajudar a proteger o DNA.
Uso incorreto de CIAP ou ineficiente inativação.
Quantidade excessiva de DNA
Digestão incompleta do DNA. Um vetor mal digerido pode levar a um elevado número de plasmídeos vazios. Isto pode ser avaliado fazendo-se uma ligação controlo, sem fragmento a inserir. Quando o fragmento é mal digerido, também levará a ineficiência da ligação. Quando se digerir o produto de PCR, deve-se garantir que sejam adicionadas bases azotadas extra nos primers usados na amplificação, para que as enzimas de restrição consigam associar-se melhor às extremidades do fragmento (as enzimas de restrição precisam de um número mínimo de bases para digerem eficientemente o produto de PCR – informação disponível no catálogo da New England Biolabs).
Ligação incompleta. Extremidades abruptas e algumas extremidades coesivas que têm baixa Tm requerem mais ligase e maior tempo de incubação.
Proteína expressa pelo gene ligado é tóxica para as células.
Sequências homólogas entre o plasmídeo e o fragmento a inserir, resultando em deleções.
Imagens: Imagem de Destaque | Extremidades | Ligação
{kind=link}
{kind=link}
{kind=link}