
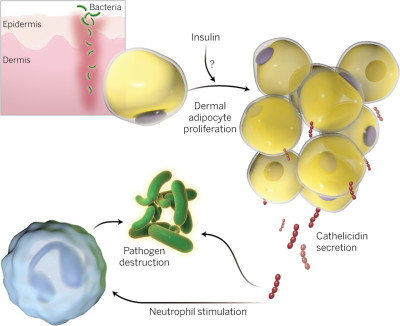
A pele é o maior órgão do nosso organismo sendo a primeira barreira contra a entrada de patogéneos nos nossos tecidos. É composta pela sua camada externa, a epiderme e pela camada mais interna, a derme que é inundada por tecido vascular, fibroblastos, fagócitos, linfócitos e tecido adiposo. A sua rutura pode levar ao estabelecimento de uma infeção bacteriana. Quando tal acontece várias células presentes quer na derme quer na epiderme limitam a colonização das bactérias. É por exemplo o caso dos queratinócitos presentes epiderme. Estes produzem péptidos antimicrobianos que destroem os patogéneos e juntamente com fagócitos na derme levam à eliminação dos mesmos. No entanto o papel dos adipócitos na defesa da pele de um indivíduo só agora emergiu. Num artigo publicado na revista Science na passada semana, Zhang et al. demonstraram que os adipócitos presentes na pele participam diretamente na imunidade inata contra a bactéria Staphylococcus aureus.
S. aureus, bactéria comensal comummente encontrada na nossa pele é responsável por mais mortes nos EUA que qualquer outro patogéneo, em particular a estirpe resistente a meticilina (MRSA). A suscetibilidade a infeções cutâneas e pulmonares por S. aureus tem sido associada a uma diminuição da produção de citocinas nos pulmões e pele respetivamente. Estas são responsáveis por regular a produção de péptidos antimicrobianos pelo epitélio e pelas células do sistema imunitário. S. aureus leva também à produção de interleucina-6 pelos adipócitos, uma citocina que estimula a produção de hepcidina, uma proteína que liga ferro e possui um efeito bacteriostático. Indicação de que os adipócitos possuem um papel na defesa do indivíduo contra este patogéneo.
Uma descontinuidade na epiderme devida por exemplo a um corte pode levar à infeção da derme por S. aureus resultando em inflamação (celulite e fasciite). De forma a simularem a instalação da infecção, Zhang et al. usaram injeções subcutâneas de MRSA em ratos. A infeção leva ao recrutamento de células mieloides, linfócitos e mastócitos resultando na eliminação da bactéria com consequente defesa do organismo. No entanto a derme possui também tecido conjuntivo onde se encontram fibroblastos e adipócitos. Os autores notaram que a infeção por MRSA levava a um aumento do tecido adiposo na derme, em parte devido à hipertrofia e proliferação dos adipócitos. Ocorre ainda adipogénese devido ao fator de transcrição Zinc Finger Protein 423 (ZFP423) cuja expressão controla a expressão de um outro fator de transcrição denominado Peroxissome Proliferator-Activated Recetor Gamma (PPAR-γ). Utilizando ratos mutados para ZFP423 ou tratando ratos saudáveis com inibidores de PPAR-γ, Zhang et al. revelaram a necessidade desses fatores na expansão do tecido adiposo na derme em resposta a uma infeção por MRSA. Ao impedir a adipogénese na pele dos ratos há diminuição nas defesas do organismo contra infeções por MRSA.
Catelicidina, um péptido antimicrobiano com atividade antiestafilocócica. Ao demonstrar que uma linha de células adiposas de ratinho assim como adipócitos humanos produzem este péptido quando cultivadas num meio com S. aureus, Zhang et al. sugeriram que as células adiposas são capazes de detetar diretamente o patogéneo. Verificou-se ainda que adipócitos de ratos com deficiência em catelicidina não foram capazes de controlar o crescimento de S. aureus. Animais com uma adipogénese deficiente possuem uma baixa produção de catelicidina após infeção com S. aureus sendo tão suscetíveis a infeção como os ratos com deficiência em catelicidina. Verificaram também que a inibição de PPRA-γ em ratos com deficiência em catelicidina não piora a infeção sugerindo que a maior proteína antiestafilocócica controlada pela adipogénese é a catelicidina.
Conclui-se então que para além do seu papel no crescimento e metabolismo, os adipócitos comportam também um importante papel no controlo de infeções nos tecidos moles. De uma perspetiva evolucionária faz todo o sentido uma vez que providencia ao indivíduo uma camada protetora adicional contra uma ferida traumática na epiderme. Contudo existe uma quantidade saudável de gordura. Zhang et al. pensaram nisto ao estudar quais os efeitos de uma dieta rica em gordura. Denotaram que a indução da adipogénese em ratos através de uma dieta rica em gordura também leva a um aumento da produção de catelicidina. No entanto ratos que possuam uma mutação para o recetor de leptina – uma hormona produzida pelos adipócitos que suprime a ingestão de alimentos – aumentam de peso e desenvolvem diabetes tipo II, mas são mais suscetíveis a infeções por S. aureus apesar de possuírem um maior número de células adiposas. De igual forma, nos humanos, a obesidade está associada com um risco aumentado de infeções cutâneas e nos tecidos moles. Uma possível explicação para esta discrepância é que a resistência à insulina ou outros aspetos de síndrome metabólica perturbem a via infeção-adipogénese-catelicidina identificada por Zhang et al. Assim sendo a sinalização por hormonas produzidas nos adipócitos que controlam o gasto energético podem influenciar a expressão de catelicidina. Este péptido é clivado pós-traducionalmente na sua forma ativa, um processo que pode ser influenciado pela obesidade e síndrome metabólica.
O mecanismo por detrás do reconhecimento da bactéria S. aureus pelos adipócitos é ainda incerto, contudo é provável que envolva o toll-like receptor 2 (TLR2). Sabe-se que os adipócitos expressam vários membros da família toll-like receptor incluindo o TLR2 responsáveis pelo reconhecimento de lipopéptidos produzidos pela bactéria. Pode ser esta a via que controla a produção de catelicidina, TLR2-ZFP423-PPAR-γ-catelicidina. Esta pode ser estimulada farmacologicamente por agonistas de PPAR-γ, aumentando assim a resistência do indivíduo contra infeção, especialmente em indivíduos suscetíveis como é o caso de indivíduos com diabetes ou síndrome metabólica.
Science 2 January 2015: 347 (6217), 26–27. [DOI:10.1126/science.aaa4567]
Science 2 January 2015: 347 (6217), 67–71. [DOI:10.1126/science.1260972]